
|
| |||
|
Разделы
Главная
Сапромат
Моделирование
Взаимодействие
Методы
Инновации
Индукция
Исследования
Факторизация
Частоты
Популярное
Как составляется проект слаботочных сетей?
Как защитить объект?
Слаботочные системы в проекте «Умный дом»
Какой дом надежнее: каркасный или брусовой?
Как правильно создавать слаботочные системы?
Что такое энергоэффективные дома?
|
Главная » Зависимость эндотелий Зависимость эндотелийзависимой поток-ндуцируемой реактивности артерий от величины трансмурального давления Филатова О.В. fkon-filatov@yandex.ru) Алтайский государственный университет Введение С начала 80-х годов пристальное внимание исследователей, работающих в области сердечно-сосудистой физиологии привлекает феномен эндотелийзависимой поток-индуцируемой (ЭЗПИ) регуляции артериальных сосудов. В ряде работ показано, что внутренняя выстилка артерий эндотелий способен чувствовать напряжение сдвига ближайшего к нему слоя жидкости [3, 4, 5, 10, 12, 22], в ответ на увеличения скорости потока выделять эндотелиальный релаксирующий фактор [15], которым является оксид азота NO [17, 19, 20]. В настоящий момент большое внимание уделяется молекулярно-клеточным механизмам ЭЗПИ реактивности артерий [2, 17, 19, 20, 21], установлению функционального значения этого вида регуляции диаметра артериальных сосудов [7, 8, 9]. Как правило, поток-зависимые реакции кровеносных сосудов исследовались в условиях трансмурального давления 100 мм рт.ст. [1, 3, 4, 5, 10, 12, 16, 22]. Неизвестно, каким образом будет проявляться ЭЗПИ реакция в широком диапазоне артериального давления. Целью данного исследования было изучение эндотелий-зависимой реакции сонных артерий кроликов на увеличение скорости кровотока в зависимости от величины трансмурального давления Методика Острые опыты проводились на кроликах массой 3,5 - 4 кг, наркотизированных кетамином (13 мг/кг внутривенно, премедикация аминазином). Сонная артерия выделялась на протяжении 4 см, артерию щитовидной железы перевязывали, исследуемый сосуд надрезали, канюлировали, сохраняя длину равной in situ. Перфузия сосуда осуществлялась по методике описанной ранее [11]. После 60 минутного периода стабилизации артерию перфузировали цельной кровью с добавлением гепарина, непульсирующим потоком с объемной скоростью 7,5 мл/мин при трансмуральном давлении 100 мм рт. ст. Объемную скорость потока жидкости увеличивали с шагом 7,5 мл/мин. Поток-зависимую вазодилатацию изучали до момента прекращения увеличения диаметра артерии в ответ на увеличение скорости потока (рис. 1), оценивали по показателю прироста диаметра, выраженному в процентах AD%. Для изучения сократимости гладких мышц (ГМ) артерий в перфузируемом сосуде в условиях постоянной объемной скорости потока 7,5 мл/мин повышали давление с шагом 10 мм рт. ст. при обработке артерий норадреналином (0,8 мкг/мл) и папаверином (10 мкг/мл). Данные представлены в виде среднего ± ошибка среднего. Достоверность различий средних величин оценивали по t критерию Стьюдента. Типичный дилататорный ответ на повышение скорости потока сонной артерий показан на рис. 1. Рис. 1. Типичный дилататорный ответ на повышение скорости потока сонной Зависимость диаметра сонной артерии от объемной скорости потока имеет нелинейный S-образный характер, выходит на плато при величине объемной скорости 52,5 мл/мин (рис. 2). Скорость кровотока в покое для сонных артерий кролика составляет 30 мл/мин в покое [Langille L. B. 1986]. Эндотелийзависимая поток-индуцируемая реакция магистральных артерий в ответ на восьмикратное увеличение объемной скорости потока (с 7,5 до 60 мл/мин) имеет максимум при 20 мм рт. ст. (30,6±1,65 %) и прогрессивно снижается до 0% при 250 мм рт. ст. (рис. 3). Прогрессивное снижение потоковой реакции с повышением давления может быть обусловлено рядом причин: снижением реактивности эндотелия, который претерпевает определенные механические трансформации, изменением степени взаимоперекрытия эндотелиальных клеток, изменением протяженности диффузионных Результаты исследования и их обсуждение 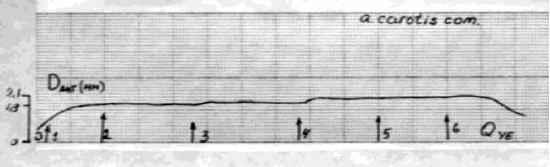 артерии. 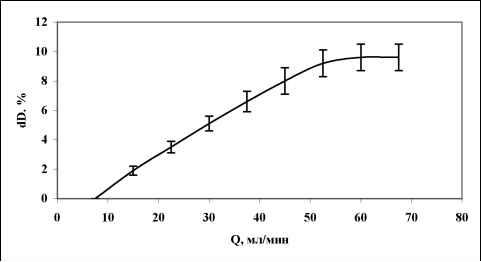 Рис. 2. Зависимость прироста диаметра общей сонной артерии кролика вызванного увеличением объемной скорости кровотока при величине трансмурального давления 100 мм рт. ст. (n=17). 35 -, 30 25 -20 15 10 0 50 100 150 200 250 300 Р, мм рт. ст. Рис. 3. Зависимость прироста диаметра, вызванного увеличением объемной скорости кровотока, от величины трансмурального давления. (n=17). При растяжении кровеносного сосуда изменяется соотношение длина-сила для ГМ. При увеличении диаметра кровеносного сосуда меняется относительная толщина сосудистой стенки, в соответствии с законом Лапласа изменяются условия передачи усилия, развиваемого ГМ. 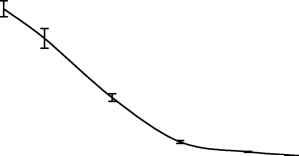 Помимо перечисленного не следует забывать, что по мере увеличения давления увеличивается напряжение пассивного скелетона кровеносного сосуда, возрастает жесткость системы кровь - сосудистая стенка, то есть режим работы ГМ последовательно трансформируется от изотонического при давлении близком к нулю - до изометрического при давлении близком к 250 мм рт. ст. Следствием повышения трансмурального давления является увеличение диаметра, т.е. увеличивается площадь поперечного сечения сосуда, а это может вести к снижению линейной скорости потока и уменьшению напряжения сдвига. Снижение выделения ЭРФ в этих экспериментах может быть следствием уменьшения напряжения сдвига, а не повышения давления. На рисунке 4 изображена усредненная зависимость диаметра от давления (D-P) общей сонной артерии кролика при обработке сосудов норадреналином и папаверином. 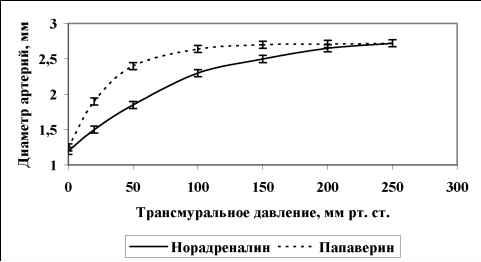 Рис. 4 Зависимость диаметра общей сонной артерии кролика от величины трансмурального давления (n=21). Зависимость активности ГМ от давления, отражающая соотношение длина - сила для гладких миоцитов, изученная по способу, предложенному Dobrin P. B. и Rovick (1969), и полученная вычитанием кривой нагружения D-P, обработанной норадреналином, из соответствующей кривой сосуда, обработанного папаверином, по своей сути, представляет зависимость Франка-Старлинга для гладких мышц. Кривая демонстрирует увеличение активности гладких миоцитов до величины давления 50 мм рт. ст., затем - прогрессивное снижение активности с увеличением трансмурального давления до 250 мм рт. ст. (рис. 5). Сопоставление зависимости S 0,5 1 я j 0,4 о о я с 0,3 -j 0,2 0,1 -i 0 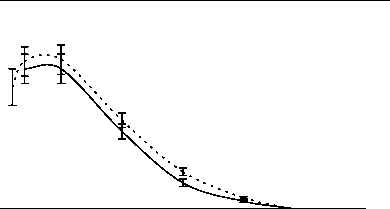 0 50 100 150 200 250 Трансмуральное давление, мм рт. ст. Активное сокращение Прирост диаметра Рис. 5. Зависимость активного сокращения ГМ (n=21) и прироста диаметра в ответ на увеличение объемной скорости кровотока (n=17) от величины трансмурального давления. По мере увеличения давления режим работы ГМ меняется от изотонического до изометрического. Исходный диаметр артерии определяет ее резервные возможности к релаксации. В диапазоне больших давлений ГМ предельно растянута, находится в изометрическом режиме, ее релаксационные возможности практически исчерпаны, поток-зависимая реакция артерии снижается до 0%. Выявленная зависимость согласуется с литературными данными, посвященными исследованию зависимости силы ГМ от длины и поперечного сечения. P. H. Cooke и F. S. Fay (1972), изучая распределение на поперечном срезе толстых и тонких филаментов растянутых и нерастянутых мышечных клеток тонкого кишечника, показали, что филаменты сократительного аппарата ГМ образуют трехмерную сеть, в углах которой находятся плотные тельца и миофибриллы диаметром 100 А. При растяжении гладкомышечной клетки плотные тельца и филаменты диаметром 100 А скапливаются в центральной части клетки, сеть вытягивается и может влиять на возможность активного сокращения от давления с кривой прироста диаметра в широком диапазоне трансмурального давления позволяет утверждать, что снижение прироста диаметра с увеличением давления обусловлено изменением режима работы гладких мышц. сократительного аппарата эффективно взаимодействовать в растянутых фибриллах. Зависимость развиваемой силы от длины определяется направленностью толстых и тонких филаментов и степени их перекрытия в различной степени растяжения. Резюмируя можно сказать - эндотелийзависимая поток-индуцируемая реакция магистральных артерий модулируется величиной трансмурального давления, имеет максимум при 20 мм рт. ст. и прогрессивно снижается до 0% при 250 мм рт. ст., что обусловлено изменением режима работы гладкой мускулатуры от изотонического к изометрическому. Литература 1. Балашов С. А., Мелькумянц A. M. Изменение тонуса сонных артерий кошек в ответ на изменение скорости кровотока Бюллетень экспериментальной биологии и медицины. - 1984. - N 11. - С. 515-517. 2. Корж А. Н. Современные представления о структуре, функции и биологической роли сосудистого эндотелия Междунар. мед. ж. (Харьков). - 2003. - Т. 9, № 1. -С. 130-134. 3. Мелькумянц A. M., Веселова Е.С. Чувствительность артерий к скорости тока и вязкости крови Тр. первого всесоюзного биофизического съезда. - М., 1982. - С. 56-60. 4. Мелькумянц A. M., Балашов С. А. Скорость кровотока постоянно действующий фактор дилатации артерий Бюл. экспериментальной биологии и медицины. -1985, М 1, С.7-9. 5. Мелькумянц A. M., Балашов С. A. Обусловленная эндотелием регуляция артерий соответственно напряжению сдвига Роль эндотелия в физиологии и патологии сосудов. - М., 1989. - Т. 38. - С. 27-60. 6. Мелькумянц А. М., Балашов С. А., Картамышев С. П. Антиконстрикторный эффект чувствительности эндотелия к напряжению сдвига Физиологический журнал им. Сеченова. - 1992. - Т. 82, № 4. - С. 93-101. 7. Мелькумянц А. М., Балашов С. А., Картамышев С. П. Роль механочувствительности эндотелия в ослаблении констрикторных реакций сосудистого русла Российский физиологический журнал им. Сеченова. - 2004. - Т. 90, № 6. - С. 693-704. 8. Осадчий Л. И., Балуева Т. В., Сергеев И. В. Эндотелийзависимый механизм формирования реакций системной гемодинамики Рос. физиол. Журн. Им. И. М. Сеченова. - 2003. - Т. 89, № 7.- С. 810-816. 9. Осадчий Л. И., Балуева Т. В., Сергеев И. В. NO-зависимый механизм реакций системной гемодинамики Рос. физиол. Журн. Им. И. М. Сеченова. - 2004. - Т. 90, № 8.- С. 490-491. 10. Смешко В., Хаютин В. М., Герова М., Геро Я., Рогоза А. Н. Чувствительность малой артерии мышечного типа к скорости кровотока - реакции самоприспособления просвета артерии Физиологический журнал СССР. - 1979. -Т. 65, № 2. - С. 291-298. 11. Филатова О. В., Киселев В.Д. Эндотелий-зависимая регуляция диаметра сонных артерий потоком в разных возрастных группах кроликов Физиологический журнал им. М.И. Сеченова, 1998, Т. 84, №5-6, с. 500-506. 12. Хаютин В. М. Регуляция просвета артерий, определяемая чувствительностью эндотелия к скорости течения и вязкости крови Вестник АМН СССР. - 1987. - N 6. - С. 89-95. 13. Cooke P. H., Fay F. S., Correlation between fiber length, ultrastructure and the length-tension relationship of mammalian smooth muscle J. Cell. Biol. - 1972. - V. 52, N 1. -P. 105-116. 14. Dobrin P. В., Rovick A. A. Influence of vascular smooth muscle on contactile mechanics and elasticiti of arteries Amer. J. Physiol. - 1969. - V. 217, № 6. - P.1644-1651. 15. Furchgott R.F. The role of endothelium in the responsis of vascular smootz muscle to drugs Annu Rev-Pharmacol. Toxicol, 1984, V 24- P. 175 - 197. 16. Gerova H., Smiesko V., Gero J., Batza E. Dilatation of conduit coronary artery induced by high wlood flow Physiol. Bohemoslov. - 1983. - V. 32, N 1. - P. 55-63. 17. Griffith. M. Aminoguanimne selectively inhibits inducible nitric oxide synthase Br. J. Pharmacology, 1993, V. 110- P. 226-230. 18. Langille L. B., ODonnell F. Redustion by chronic decreases in blood flow are endothelium dependent Science. - 1986. V. 231. - P 86-89. 19. Moncada S., Herman A.G., Vanhoute P. Endothelium-derived relaxing factor is identified as nitric oxide Trends Pharmaeol. Sci. - 1987. - V. 8, № 10. - P. 365-368. 20. Palmer R. M. J., Ferrige A. G., Moncada S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor Nature. - 1987. - V. 327. - P. 524-526. 21. Sausbier M., Schubert R., Voigt V., Hirneiss C., Pfcifcr A., Korth M., Kleppisch T., Ruth P., Hofmann F. Mechanisms of NO/cGMP-dependent vasorelaxation. Circ Res. - 2000. - V. 87, № 9. - C. 825-830. 22. Smiesko V., Kozik J., Dolezel S. Role of endothelium in the control of arterial diameter by blood flow Bitood Vessels. - 1985. - V. 22. - P. 247-251. |
|
| |||