
|
| |||
|
Разделы
Главная
Сапромат
Моделирование
Взаимодействие
Методы
Инновации
Индукция
Исследования
Факторизация
Частоты
Популярное
Как составляется проект слаботочных сетей?
Как защитить объект?
Слаботочные системы в проекте «Умный дом»
Какой дом надежнее: каркасный или брусовой?
Как правильно создавать слаботочные системы?
Что такое энергоэффективные дома?
|
Главная » Связанность 1 2 Связанные с событиями потенциалы мозга при восприятии длительности стимулов. Сообщение II: короткие слуховые сигналы Ходанович М.Ю.(Mhodanovich@/mail.ru) (2), Бушов Ю.В (1), Иванов А.С.(1), Вячистая Ю.В.(2) (1)Томский государственный университет (2) НИИ биологии и биофизики Согласно литературным данным [1], минимально возможный для различения интервал времени составляет 50-100 мс - так называемая психологическая единица времени. С другой стороны убедительно показано, что компонент негативность рассогласования (НР) связанных с событиями потенциалов мозга (ССПМ) появляется даже тогда, когда различия физических характеристик звуковых стимулов не детектируются испытуемым на уровне сознания [2]. Установлено также, что этот компонент не зависит от наличия произвольного внимания, а генерируется мозговыми процессами автоматически при различии физических характеристик в последовательно предъявляемых стимулах [3]. Вероятно, регистрируя эту волну при разной величине отклонения длительности девиантного стимула от стандартного, можно определить минимальный интервал времени, детектируемый на уровне автоматической обработки информации и не достигающий уровня сознания. Несмотря на большое количество исследований, посвященных НР, зависимость этого компонента от величины различий между предъявляемыми стимулами ( стимульного отклонения - СО) изучена недостаточно. В целом литературные данные свидетельствуют о том, что амплитуда НР увеличивается с ростом СО [4]. Однако, есть также работы, в которых небольшое, недетектируемое СО вызывало НР примерно той же самой величины, что и детектируемое [5]. Относительно небольшое число работ использует в качестве СО длительность стимула [7, 8, 9]. При этом, влияние самой длительности, независимо от других характеристик стимула и экспериментальных условий, как правило, не учитывается. С другой стороны, известно, что такие компоненты ССПМ, как устойчивый потенциал, появляются том же участке, что и НР [10], существенно зависят от длительности стимула [14], и поэтому могут перекрываться c НР. Исследование этой зависимости необходимо как для понимания нейрофизиологических основ различения интервалов времени, так и для исследования НР, которая на сегодняшний день признана объективным коррелятом автоматической обработки информации о параметрах стимула. Компонент процессная негативность (ПН), являющийся коррелятом селективного внимания и связанный со сличением стимула с эталоном, достаточно хорошо изучен при различении частоты, громкости стимулов [5]. В значительно меньшей степени этот компонент изучался в ситуации различения стимулов по длительности. Таким образом, целью данного исследования стало изучение слуховых ССПМ при восприятии длительности стимулов в условиях активного внимания к этим стимулам и в его отсутствие. Методы исследования Объект исследования. В исследованиях участвовали 22 практически здоровых добровольца (9 мужчин, 13 женщин) в возрасте от 18 до 24 лет, студенты томских вузов. Работа испытуемых оплачивалась. Стимулы и задача. В качестве звуковых стимулов использовались тоны интенсивностью 60 дБ, частотой 2000 Гц, генерируемые компьютером, которые предъявлялись бинаурально через динамики. В ходе эксперимента испытуемым предъявлялись частые стандартные и редкие девиантные стимулы с вероятностью предъявления соответственно 80 и 20%. Длительность сигналов задавалась с точностью до 1 мс. В качестве девиантных использовали стимулы длительностью 200 мс, в качестве стандартных - в разных блоках стимулы длительностью 400, 300, 250, 220, 180, 150 и 100 мс (режимы 400-200 , 300-200 , 250-200 , 220-200 , 180-200 , 150-200 , 100-200 ). Для создания условий отсутствия внимания к длительности стимула испытуемому предлагалось чтение научно-популярного журнала ( Наука и Религия ). Для привлечения внимания к длительности стимула испытуемому сначала предъявляли для запоминания 5 стимулов эталонной длительности (девиантный стимул), а затем в псевдослучайном порядке следовали стандартные и девиантные стимулы с межстимульным интервалом 1000 мс. Если испытуемый узнавал девиантный стимул, он должен был нажать на клавишу пробел . В течение выполнения задания эталонные стимулы предъявлялись несколько раз для напоминания (5 блоков стимулов по 50 стимулов в каждом блоке, всего 250 стимулов для каждой из длительностей). На случайную последовательность стимулов накладывались следующие ограничения: целевой (девиантный) стимул не был первым в блоке и между девиантными стимулами был хотя бы один стандартный. В этих условиях по литературным данным (Squires K.C., Wickens C., Squires N.K., Donchin E., 1976) наиболее выражен компонент негативность рассогласования . По результатам теста подсчитывали процент правильных и ошибочных реакций испытуемого, а также среднее время реагирования. Регистрация электрофизиологических показателей. Параллельно с предъявлением стимулов регистрировали ЭЭГ по стандартной системе 10-20% в отведениях F3, Fz, F4, С3, Cz, С4, Результаты исследований Точность различения стимулов и время реакции. Из таблицы 2 следует, что при различиях стандартного и девиантного стимулов на 100 и более мс, целевой стимул 200 мс Р3, Pz, Р4, T3, T4, T5, T6, О1, О2 с помощью 16-ти канального энцефалографа ЭЭГ-16 фирмы Medicor в частотном диапазоне от 0.23 до 2000 Гц. Объединенный референтный электрод устанавливался на мочки левого и правого уха, а земляной фиксировался на запястье испытуемого. При регистрации ЭЭГ использовали клеющиеся электроды фирмы Nicolet . С целью устранения артефактов, связанных с движениями глаз и электродермальной активностью, параллельно с записью ЭЭГ регистрировали КГР (по методу Тарханова) и ЭОГ. При вводе аналоговых сигналов в ЭВМ шаг дискретизации составил 1 мс. Выделение и фильтрацию связанных с событиями потенциалов мозга (ССПМ) проводили способом, описанным в Сообщении I. Эпоха анализа, включая фоновый фрагмент 100 мс, составляла 600 мс. Статистическая обработка. Математическая обработка данных проводилась с помощью пакета прикладных программ для статистического анализа Statistica 6.0, а также самостоятельно разработанных программ для предварительной обработки данных на языке Delphi 5.0 и Visual Basic for Applications для Statistica 6.0. Для оценки распознавания стимулов использовали показатель эффективность узнавания (ЭУ) (Сообщение I), который сравнивали с пороговой ЭУ с помощью Т-критерия и критерия Хи-квадрат. Для анализа ЭУ и времени реакции в сериях с разными длительностями стимулов использовался также непараметрический ранговый критерий Вилкоксона. Различия между ССПМ на разные типы стимулов оценивали двумя способами. 1) Поточечно для каждого отведения с помощью критерия Вилкоксона для сравнения двух типов стимулов и рангового дисперсионного анализа Фридмана для сравнения нескольких типов стимулов; 2) Выделялись следующие компоненты ССПМ: НР/№Ь в ССПМ на девиантный стимул и УП в ССПМ на стандартный стимул как максимальная негативность на участке 200-400 мс от начала стимула. Для оценки влияния факторов длительности стимула, величины и знака стимульного отклонения использовался дисперсионный анализ ANOVA c повторными измерениями: модель с двумя уровнями (длительность стимула и область отведения) для оценки влияния длительности на амплитуду УП и модель с тремя уровнями (знак отклонения, величина отклонения и область отведения) для оценки влияния параметров отклонения стимула на амплитуду НР/№Ь. Все переменные, для который применялся Т-тест и дисперсионный анализ, исследовались на нормальность распределения. Для числа степеней свободы больше двух проводилась коррекция Greenhouse-Geisser.
Примечание. Звездочками отмечены значимые отличия частоты правильных ответов от их ожидаемой частоты в ситуации когда испытуемый не различает стимулы по длительности (критерий Хи-квадрат: * - p<0.05, ** - p<0.01, *** - p<0.001, Т-критерий: # - p<0.05, ## - p<0.01, ### - p<0.001). Дисперсионный анализ выявил значимое влияние величины СО на точность распознавания (F(2, 28)=17.884, p=0.00001), влияние знака этого отклонения на точность оказалось незначимым (F(1, 14)=1.6492, p=0.2). Время реакции отрицательно коррелировало с ЭУ звуковых стимулов (r=-0.47-0.81, p<0.05-0.01). Различия ССПМ на стандартный и девиантный стимулы при отсутствии внимания к звуковым стимулам. В условиях отвлечения внимания в сериях с редким увеличением длительности ( 100- 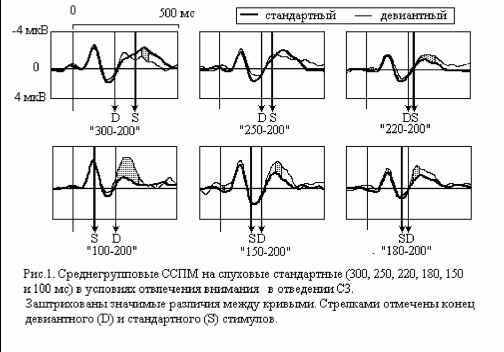 200 , 150-200 и 180-200 ) на участке 120-350 мс от начала стимула наблюдается статистически значимый негативный сдвиг потенциала на девиантный стимул (рис.1). Этот сдвиг хорошо распознавался испытуемыми (ЭУ составляет 77-97%), при различиях на 20 мс - не распознавался. При различиях в длительности на 50 мс согласно Т-критерию целевой стимул распознавался, согласно более жесткому непараметрическому критерию Хи-квадрат - не распознавался испытуемыми. Установлено, что во всех сериях, где интервал времени задавалсяся звуковыми стимулами (кроме 150-200 ), точность распознавания оказалась существенно выше (p<0.05-0.001) по сравнению с точностью узнавания тех же интервалов, задаваемых зрительными стимулами (Сообщение I). Таблица 1. Точность различения стандартного (в разных сериях 400, 300, 250, 220, 180, 150 и 100 мс) и девиантного (200 мс) стимулов и среднее время реакции при опознании звуковых стимулов. 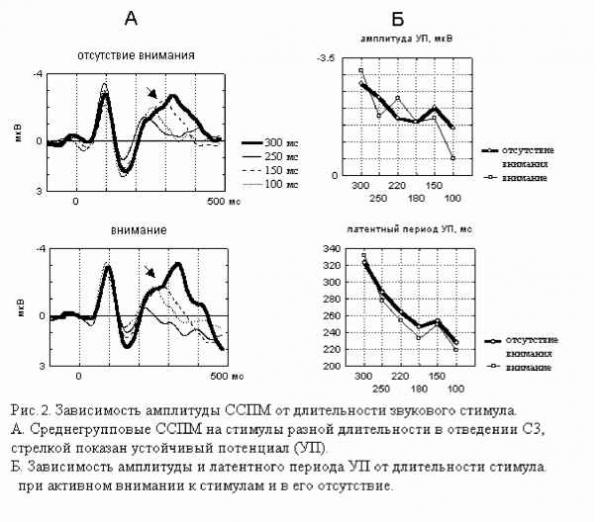 начинается с 120-140 мс в серии 100-200 , с 160 - 180 мс в серии 150-200 и с 220 мс в серии 180-200 , достигает максимума в отведениях F3 и С3 на 240-270 мс и продолжается до 300-350 мс от начала стимула. Максимальная амплитуда этой разностной волны (РВ) составила 4 мкВ в сериях 100-200 , 1.5-2 мкВ в серии 150-200 и 1 мкВ в серии 180-200 . Мы полагаем, что эти волновые различия связаны с появлением компонента негативность рассогласования (НР)[4]. В серии 100-200 в ССПМ на девиантный стимул на участке от 350 до 500 мс от начала стимула также наблюдается значимая позитивная волна с амплитудой 0.5-1 мкВ. По нашему мнению, этот сдвиг соответствует компоненту Р3а, связанному в включением непроизвольного внимания [6]. В сериях, где длительность стандартного стимула (300, 250 и 220 мс) превышала длительность девиантного (200 мс), результаты отличаются от ожидаемых и поэтому представляют несомненный интерес (рис.1). Ожидалось, что в этих сериях РВ также будет негативной, но несколько большей пиковой латентности. Однако, в сериях 300-200 и 250-200 на участке 250-500 мс от начала стимула сдвиг потенциала на девиантный стимул, напротив, был позитивным (1.5-2 мкВ для серий 300-200 и 0.5-1 мкВ для серий 250-200 ). Зависимость характеристик ССПМ от длительности стимула. При сравнения ССПМ на стандартные стимулы разной длительности (рис.3) с помощью дисперсионного анализа обнаружены близкое к значимому влияние фактора длительности стимула (F(5, 45)=2.0, p=0.10402) и значимое взаимодействие факторов длительности стимула и области отведения (F(70, 630)=1.4910, p=0.008) на негативный компонент с ЛП 200-400 мс от начала стимула. Эффект проявился в том, что во фронтальных, центральных и париетальных отведениях негативный сдвиг потенциала увеличивается пропорционально длительности стимула. Различия между сериями ССПМ на стимулы длительностью 100 и 400 мс составляли до 4 мкВ. Вероятно, этот компонент представляет собой устойчивый потенциал (УП) [9]. Поточечный непараметрический дисперсионный анализ, проведенный отдельно для каждого отведения показал, что амплитуда и латентность УП достоверно увеличиваются с ростом длительности стимула в фронтальных, центральных, париетальных и передневисочных отведениях (p<0.001, критерий Фридмана) как при активном внимании, так и в его отсутствие. Влияние внимания на амплитуду УП оказалось незначимым. Зависимость характеристик ССПМ на девиантный стимул от величины СО. Поскольку сравнение разностных ССПМ не дает четкого представления о влиянии величины СО на амплитуду НР из-за присутствия УП, был проведен сравнительный анализ ССПМ на девиантный стимул в сериях с отвлечением внимания. При сравнении НР/№Ь с помощью дисперсионного анализа обнаружен главный эффект фактора величины СО на амплитуду этого компонента, связанный с увеличением амплитуды с ростом СО (F(2,18) = 4.6141, p=0.02) (рис.3). Обнаружен также значимый главный эффект фактора знака отклонения на амплитуду НР/№Ь (F(1,9) = 14.136, p=0.005), связанный с тем, что при редком увеличении длительности стимула амплитуда НР/№Ь была больше, чем при редком уменьшении. Зарегистрировано также значимое взаимодействие факторов знака (F(14, 126)=5.4268, p=0.00000) и величины отклонения (F(28, 252)=2.5401, p=0.00007) с фактором области отведения. Оба эти эффекта связаны с тем, что различия между сериями были наиболее выражены во фронтальных и центральных отведениях. Обнаружены значимые положительные корреляционные связи амплитуды НР/№Ь с ЭУ для серии 250-200 , 220-200 и 180-200 (r=0.64-0.82, p<0.05-0.001). Различия ССПМ, связанные с активным вниманием к длительности стимула. Начиная с 350 мс от начала стимула в сериях 100-200 и 150-200 и с 400 мс в сериях 300-200 и 250200 в ССПМ на целевой девиантный стимул во всех отведениях наблюдается значимый позитивный сдвиг потенциала с максимумом 5 мкВ на 380-410 мс в отведениях С3 и Pz. Вероятно, этот компонент соответствует P3b на целевой стимул, связанный с опознанием стимула [4]. Известно, что компонент процессная негативность (ПН) может быть получен вычитанием из ССПМ на целевой релевантный стимул ССПМ на такой же стимул в условиях игнорирования [5], при этом выявляются изменения ССПМ, связанные с селективным вниманием и не связанные с А уменьшение длительности ц О 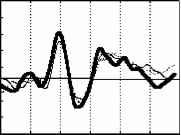 300-200 250-200 220-200 Зависимость амплитуды НР от знака стимульного отклонения (F(1, 9)=14.13б. р=0.005) 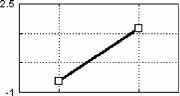 уменьшение увеличение 500 мс 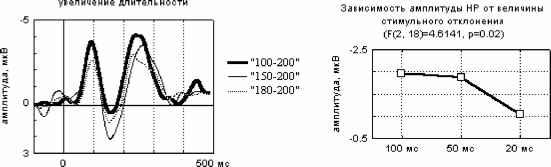 Рис. 3. Зависимость амплитуды НР от величины и знака стимульного отклонения в условиях отвлечения внимания. А. Среднегрупповые ССПМ на девиантный стимул 200 мс в отведении СЗ, вверху -для серий с редким уменьшением длительности стимула ( 300-200 , 250-200 , 220-200 ), внизу - для серий с увеличением длительности стимула ( 100-200 , 150-200 , 180-200 ). Б. Зависимость амплитуды HP/N2b от знака н величины стимульного отклонения. игнорируемым стимулами обнаружено значимое влияние фактора межполушарной асимметрии, связанное с преобладанием ПН в левом полушарии (F(1, 9)=5.9391, p=0.03). В сериях, где целевой стимул был короче игнорируемого, статистически значимая ПН наблюдалась только в серии 300- другими атрибутами стимула. При этом различия на участке до 400 мс могут быть отнесены за счет ПН, различия на участке от 400 мс - за счет РЗб на релевантный стимул [5]. Поточечное сравнение ССПМ на девиантный стимул в условиях активного внимания и в его отсутствие показало, что значимые различия, связанные с появлением негативного компонента (ПН) во фронтальных и центральных отведениях, наблюдаются во всех сериях, в которых целевым был стимул большей длительности ( 100-200 , 150-200 , 180-200 ) (рис.4). Различия начинались на 55 мс от начала стимула, достигали максимума на 160-170 мс во фронтальных отведениях и продолжались до 200-220 мс в сериях 100-200 , 180-200 и до 400 мс в серии 150-200 . При сравнении амплитуды ПН для разных временных различий между целевым и 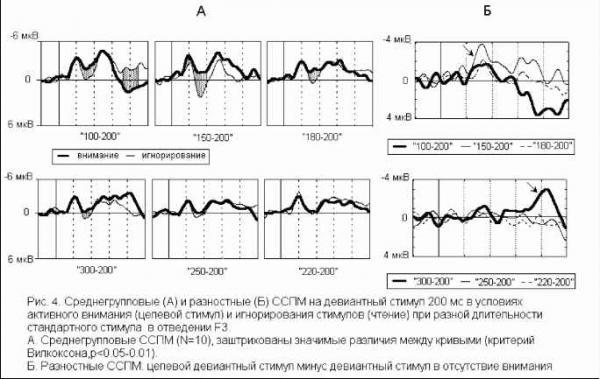 Обсуждение результатов Согласно полученным результатам, различия между стимулами в 100 мс и более мс (серии 400-200 , 300-200 и 100-200 ) распознаются испытуемыми, различия в 20 мс (серии 180200 и 220-200 ) не распознаются. При различиях в длительности между стимулами в 50 мс согласно критерию Хи-квадрат целевой стимул не опознается, согласно Т-критерию -распознается. Вероятно, различия в 50 мс являются пограничными для распознавания, что согласуется с данными литературы [ 1]. Установлено, что модальность стимула существенно влияет на точность распознавания длительности стимулов (Сообщение I). Точность распознавания слуховых стимулов оказалась существенно выше, чем зрительных. Полученные разнонаправленные корреляции со временем реакции для слуховых и зрительных стимулов, вероятно, объясняются разной ЭУ этих стимулов. Длительность слуховых стимулов в среднем распознавалась лучше, чем длительность зрительных, поэтому лучшее время реакции показали те испытуемые, для которых эта задача была проще. Для зрительных стимулов, напротив, большее время реакции показали те испытуемые, которые 200 во фронтальных отведениях на участке 190 мс от начала стимула. Дисперсионный анализ разностных ССПМ показал, что влияние фактора величины СО на амплитуду ПН незначимо. Обнаружены положительные корреляционные связи амплитуды ПН с ЭУ (r=0.59-0.73 для серий 100-200 , 150-200 , 300-200 ). вообще пытались решать эту задачу, остальные же нажимали на клавишу пробел наугад. Это предположение подтверждается тем, что наименьшее время реакции и для слуховой, и для зрительной модальностей обнаружено в тех сериях, где девиантный стимул не распознавался. Зависимость времени реакции от знака СО слуховых сигналов, вероятно, объясняется тем, предъявление коротких стимулов задает более быстрый темп деятельности. Результаты, полученные в отношении ССПМ, несколько отличаются от ожидаемых. По нашим предположениям, НР в чистом виде могла наблюдаться на временном промежутке с 200 мс от начала стимула в условиях отсутствия внимания к стимулам как негативные волновые различия между ССПМ на девиантный и стандартный стимулы. Амплитуда этих различий, согласно литературным данным [4], должна увеличиваться с ростом СО. Предположения подтвердились для серий с редким увеличением длительности стимула (серии 100-200 , 150200 , 180-200 ). В условиях отсутствия внимания НР возникает как при больших (100 мс), так и при малых (20 мс) различиях между стимулами, которые не опознаются испытуемыми. Амплитуда НР увеличивается с ростом временных различий между стимулами. Положительные волновые различия на участке 250-400 мс для серий с редким уменьшением длительности стимула ( 300200 , 250-200 и 220-200 ) отличаются от литературных данных. Так, Naatanen R., Paavilainen P., Reinikainen K. (1989) в ответ на уменьшение длительности девиантного стимула (400 мс -стандартный, 200 мс - девиантный) были получены негативные волновые различия на этом участке [8]. Причина полученной нами зависимости РВ от знака СО легко объясняется, если проанализировать зависимость ССПМ на стандартный стимул от его длительности. При увеличении длительности стимула на участке от 200 до 500 мс появляется негативный компонент, амплитуда которого увеличивается пропорционально длительности стимула. По-видимому, этот компонент представляет собой устойчивый потенциал (УП) [10]. Появление УП обычно связывают со стимулами длительностью более 600 мс [11], однако, УП зарегистрирован и на более короткие стимулы [12]. Амплитуда УП увеличивается с ростом длительности стимула [12], его ЛП составляет 150 мс, а продолжительность пропорциональна длительности стимула [13]. Вероятно, УП на стандартные стимулы большей длительности и НР на девиантный стимул меньшей длительности характеризуются одинаковой латентностью (серии 300-200 , 250-200 ). Амплитуда УП с увеличением длительности стимула, по-видимому, растет быстрее, чем НР на девиантный стимул. Поэтому в этих сериях мы наблюдали положительную РВ. Сравнение ССПМ на девиантный стимул в разных сериях показало, что амплитуда НР/№Ь зависит как от величины, так и от знака СО. Полученное нами увеличение амплитуды НР с ростом СО как при увеличении, так и при уменьшении длительности стимула, подтверждается литературными данными [7, 9]. Однако, при увеличении длительности амплитуда НР растет быстрее, чем при уменьшении длительности. При отвлечении внимания за НР при больших различиях между стимулами (100 мс) за НР следует волна Р3а, связанная с непроизвольным включением внимания, что отражается в положительных волновых различиях на участке 400-500 мс от начала стимула. В условиях активного внимания при различиях между стимулами 100 мс, которые хорошо опознаются испытуемыми (86-90% испытуемых), и 50 мс, которые распознаются 45-59% испытуемых, за НР+ПН следует компонент P3b, связанный с опознанием девиантного стимула. Компонент ПН, по данным литературы, представляет собой коррелят селективного внимания, связанный со сличением текущего стимула с эталоном [5]. Известно, что максимальная амплитуда ПН достигается при равновероятном предъявлении целевого и игнорируемого стимулов, с уменьшением вероятности предъявления целевого стимула амплитуда ПН снижается, что объясняется частичным стиранием сенсорного следа, оставленного предъявлением релевантного стимула. Так, при вероятности предъявления 1:6 амплитуда ПН снижалась в 3 раза по сравнению с равновероятным предъявлением стимулов [14]. Вероятно, этим частично объясняется отсутствие значимой ПН для серий, где релевантным был более короткий стимул. С другой стороны показано, что ПН отличается сильной межиндивидуальной вариабельностью [5], что также может объяснить отсутствие значимой ПН для этих серий. Для серий, в которых целевым был более продолжительный стимул, наблюдалось близкое к значимому влияние величины СО (р=0.1), связанное с увеличением ПН для пороговых (50 мс) значений отклонения. В этой серии ПН имела более короткий ЛП и большую продолжительность, что, по-видимому, объясняется большей сложностью различения этих стимулов [14]. Вероятно, при меньших временных различиях (20 мс) испытуемый не пытался распознавать стимулы, что проявилось в ослаблении контроля селективного внимания. Это предположение также подтверждается значительным уменьшением времени реакции в серии 220-200 (387 мс) по сравнению с серией 250-200 (513 мс). Полученные нами данные о преобладании ПН в левом полушарии не согласуются с большей частью литературных данных, связывающих ПН с правой фронтальной корой [15, 5]. Однако, в этих работах использовались другие признаки стимула: частота тона, громкость, локализация в пространстве. Возможно, ПН, связанная с анализом длительности стимула, действительно имеет другое пространственное распределение. Выводы 1. Порог различения длительности звуковых сигналов при активном внимании к этим сигналам составляет около 50 мс, а при невнимании к стимулам около 20 мс. 2. В условиях невнимания к слуховым стимулам при сравнении ССПМ на стандартные и девиантные стимулы на участке 200-400 мс от начала стимула обнаружена волна, именуемая 1 2 |
|
| |||