
|
| |||
|
Разделы
Главная
Сапромат
Моделирование
Взаимодействие
Методы
Инновации
Индукция
Исследования
Факторизация
Частоты
Популярное
Как составляется проект слаботочных сетей?
Как защитить объект?
Слаботочные системы в проекте «Умный дом»
Какой дом надежнее: каркасный или брусовой?
Как правильно создавать слаботочные системы?
Что такое энергоэффективные дома?
|
Главная » Потенциалы 1 2 Связанные с событиями потенциалы мозга при восприятии длительности стимулов. Сообщение I: короткие зрительные стимулы Ходанович М.Ю. (Mhodanovich@mail.ru) (2), Бушов Ю.В (1), Иванов А.С.(1), Вячистая Ю.В. (2) (1)Томский государственный университет (2) НИИ биологии и биофизики Установлено [1], что минимально возможный для различения интервал времени составляет 50-100 мс - так называемая психологическая единица времени. С другой стороны убедительно показано, что компонент негативность рассогласования (НР) связанных с событиями потенциалов мозга (ССПМ) появляется даже тогда, когда различия физических характеристик звуковых стимулов не детектируются испытуемым на уровне сознания [2]. Установлено также, что этот компонент не зависит от наличия произвольного внимания, а генерируется мозговыми процессами автоматически при различии физических характеристик в последовательно предъявляемых стимулах [3]. Вероятно, регистрируя эту волну при разной величине отклонения длительности девиантного стимула от стандартного, можно определить минимальный интервал времени, детектируемый на уровне автоматической обработки информации и не достигающий уровня сознания. Анализ литературы показал, что при исследовании восприятия интервалов времени авторы, как правило, используют стимулы одной модальности. Тем не менее, есть предпосылки к тому, что модальность используемого раздражителя также оказывает влияние на точность оценки интервала времени. Например, по мнению П. Фресса [4], световая субъективная секунда, в отличие от звуковой , близка к физической. В отличие от слуховой НР, условия возникновения которой достаточно хорошо изучены [5], существование зрительной НР до сих пор убедительно не доказано. Существует относительно небольшое число работ, в которых непосредственно анализируется процесс сравнения зрительной информации. Среди них есть работы, в которых этот компонент был обнаружен [6], в другой части работ его обнаружить не удалось [7]. Есть также работы, в которых компонент, подобный зрительной НР, был обнаружен только в условиях внимания к стимулам [8, 9]. Целевыми при этом являлись другие признаки стимула, поэтому указанный компонент может иметь отношение к процессам автоматической обработки информации, однако, в строгом смысле его нельзя считать аналогом слуховой НР из-за зависимости от внимания. В доступной литературе нами не обнаружены данные об изменениях ССПМ при отклонении зрительного стимула по длительности, а также зависимости этих изменений от величины различий между предъявляемыми стимулами ( стимульного отклонения ). Компонент процессная негативность (ПН), связанный со сличением стимула с эталоном и возникающий при активном внимании к стимулам, достаточно хорошо изучен при различении цвета, ориентации, формы зрительных стимулов [21, 5], однако не исследовался в ситуации различения стимулов по длительности. Исходя из вышеизложенного, целью данного исследования стало изучение зрительных ССПМ при восприятии длительности стимулов в условиях активного внимания к этим стимулам и в его отсутствие. Методы исследования Объект исследования. В исследованиях участвовали 13 практически здоровых добровольцев мужского пола с нормальным или скорректированным зрением в возрасте от 18 до 24 лет, студенты томских вузов. Работа испытуемых оплачивалась. Испытуемый располагался в удобном кресле в слабо освещенной звукоизолированной камере. Перед обследованием испытуемый получал инструкцию не использовать для оценки интервалов времени счет в уме, постукивание и другие аналогичные приемы. Стимулы и задача. Зрительным стимулом служил белый квадрат со стороной 2 см, появляющийся на определенное время в центре экрана монитора. В ходе эксперимента испытуемым в случайном порядке предъявлялись частые стандартные и редкие девиантные зрительные стимулы с вероятностью соответственно 80 и 20%. Для создания условий отсутствия внимания к длительности стимула предлагалось зафиксировать взор на правом верхнем углу монитора, при этом предъявляемые стимулы находились в боковом поле зрения, и производить произвольный счет в уме. Для привлечения внимания к длительности стимула испытуемому сначала предъявляли для запоминания 5 стимулов эталонной длительности (девиантный стимул), а затем в случайном порядке следовали стандартные и девиантные стимулы с межстимульным интервалом 1000 мс. В случае, если испытуемый узнавал девиантный стимул, он должен был нажать на клавишу пробел . В качестве девиантных использовали стимулы длительностью 200 мс, в качестве стандартных - в разных блоках стимулы длительностью 400, 300, 250, 150 и 100 мс (серии 400-200 , 300-200 , 250-200 , 150-200 , 100-200 ). По результатам теста подсчитывали процент правильных и ошибочных реакций испытуемого, а также среднее время реагирования. Регистрация электрофизиологических показателей. Параллельно с предъявлением стимулов регистрировали ЭЭГ, ЭОГ и КГР. ЭЭГ регистрировалась при открытых глазах и фиксированном взоре монополярно в 8 отведениях: F3, F4, С3, С4, Р3, Р4, О1 и О2 по стандартной системе 10-20 % с помощью 8-ми канального полинейрографа с постоянной времени 3 секунды, в частотном диапазоне от 0.3 до 70 Гц, входное сопротивление составляло ~100 мОм. При регистрации ЭЭГ использовали слабополяризующиеся хлорсеребрянные электроды фирмы Medicor (Венгрия). Объединенный референтный электрод устанавливался на мочки левого и правого уха испытуемого, а земляной фиксировался на подбородке. С целью устранения артефактов, связанных с движениями глаз и электродермальной активностью, параллельно с записью ЭЭГ регистрировали КГР (по методу Тарханова) и ЭОГ. Все указанные показатели записывались в память компьютера через аналого-цифровой преобразователь (АЦП). При вводе аналоговых сигналов в ЭВМ шаг дискретизации составил 5.714 мс. Выделение связанных с событиями потенциалов мозга (ССПМ). При выделении ССПМ использовали лишенные артефактов участки ЭЭГ, которые выделялись визуально. Эпоха анализа, включая фоновый фрагмент 100 мс, составляла 900 мс. Исходную ЭЭГ и выделенные ССПМ фильтровали с помощью фильтра Хемминга с полосой пропускания 0-25 Гц. На основе предстимульного фрагмента длительностью 100 мс определяли базовую линию. Статистическая обработка. Математическая обработка данных проводилась с помощью пакета прикладных программ для статистического анализа Statistica 6.0, а также самостоятельно разработанных программ для предварительной обработки данных на языке Delphi 5.0 и Visual Basic for Applications для Statistica 6.0. Эффективность узнавания (ЭУ) стимулов эталонной длительности оценивали по процентному отношению числа правильно опознанных эталонных стимулов к сумме общего числа стимулов эталонной длительности с числом ошибочно опознанных сигналов. При правильном опознании всех стимулов эталонной длительности ЭУ достигает максимума - 100%. Если испытуемый не опознал ни одного эталонного стимула и при этом не разу не нажал на клавишу пробел , то ЭУ наименьшая и составляет 0%. Если испытуемый не опознает эталонные сигналы и нажимает на клавишу пробел при предъявлении стимулов любой длительности, то эффективность узнавания в этом случае стремится к величине равной процентному отношению числа эталонных стимулов к общему числу стимулов. Если полученная ЭУ значимо отличалась от пороговой ЭУ (ЭУп) при случайном нажатии на клавишу (при соотношении целевых и игнорируемых стимулов 20:80 ЭУп составляла 20%), то считали, что испытуемые распознают предъявленные им длительности стимулов. Для анализа точности различения интервалов и времени реакции в сериях с разными длительностями стимулов использовался непараметрический ранговый критерий Вилкоксона. Различия между ССПМ на разные типы стимулов также оценивали с помощью непараметрического критерия Вилкоксона, а также рангового дисперсионного анализа Фридмана Результаты исследований Точность различения стимулов и время моторной реакции. Проанализирована точность различения стандартного (400, 300, 250, 100 и150 мс) и девиантного (200 мс) стимулов и время моторной реакции в сериях с привлечением внимания к длительности стимула. Анализ ЭУ показал, что, согласно критерию Хи-квадрат ЭУ ни одной из длительностей не отличается от пороговой, согласно Т-критерию только в сериях 100-200 и 400-200 ЭУ значимо отличается от пороговой (табл. 1). Однако, и в этих сериях только у 63 и 38% испытуемых ЭУ больше 50 %. В сериях 400-200 и 100-200 ЭУ оказалась значимо более высокой, чем в остальных сериях (p<0.05 - p<0.001, критерий Вилкоксона). Следует заметить, что при одинаковых временных различиях 100 мс серии 300-200 и 100-200 значимо (р<0.05, критерий Вилкоксона) различаются между собой по частоте правильных ответов, в то время как при разных временных различиях между стимулами ( 400-200 и 100-200 , 300-200 и 150-200 ) статистически значимых различий нет. Дисперсионный анализ выявил значимое влияние разницы между длительностью девиантного и стандартного стимулов ( стимульное отклонение ) (F(1, 8)=16.751, p=0.003), близкое к значимому влияние знака этой разницы на точность распознавания (F(1, 8)=4.6117, p=0.06), а также их значимое взаимодействие (F(1, 8)=8.8701, p=0.01). Корреляционный анализ показал, что точность распознавания положительно коррелирует со временем реакции (r=0.65-0.76, p<0.05-0.01). Таблица 1. Точность различения стандартного (в разных сериях 400, 300, 250, 150 и 100 мс) и девиантного (200 мс) стимулов и среднее время реакции при распознавании длительности
Примечание. Звездочками отмечены значимые отличия частоты правильных ответов от их ожидаемой частоты в ситуации, когда испытуемый не различает стимулы по длительности (T-критерий): * - p<0.05, **- p<0,01. Время реакции значимо больше для серии 100-200 по сравнению с серией 250-200 (p<0.05, критерий Вилкоксона). для повторных измерений, поточечно для каждого отведения. Для оценки влияния длительности стимула на характеристики ССПМ амплитуда устойчивого потенциала (УП) вычислялась как максимально негативное значение на интервале 150-500 мс от начала стимула, влияние фактора длительности оценивалось с помощью рангового дисперсионного анализа Фридмана для повторных измерений. 800 мс 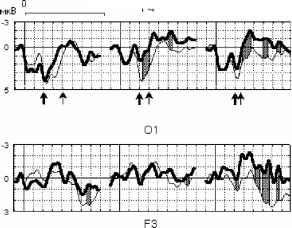 стандартный девиантный стандартный девиантный
стандартный девиантный стандартный минус девиантный 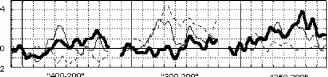 Р4 01 F3 Рис. 1. Среднегрупповые (N=10) и разностные ССПМ на стандартный (400, 300 и £50 мс) и девиантный (200 мс) стимулы в условиях отсутствия внимания к стимулу. Заштрихованы значимые различия между кривыми. 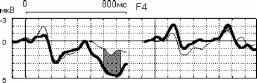 стандартный девиантный 11 и стандартный минус девиантный 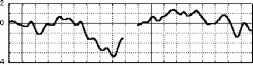 100-200 150-200 Рис 2 . Среднегрупповые (N=10) и разностные ССПМ на стандартный (100 и 150 мс) и девиантный (200 мс) стимулы в условиях отсутствия внимания к стимулу. Заштрихованы значимые различия между кривыми. 200 и 250-200 ) (рис.1). В сериях 100-200 и 150-200 подобный негативный сдвиг небольшой амплитуды (менее 1мкВ) наблюдался на участке 150 - 400 мс от начала стимула, но был статистически незначим (рис.2). В серии 250-200 максимальные значимые различия составили -3.83 мкВ (отведение О1, 500 мс от начала стимула), в серии 300-200 -2.81 мкВ (отведение Р3, 280 мс), в серии 400-200 -1.88 мкВ (отведение О1, 650 мс от начала стимула). При сравнении разностных ССПМ (девиантный стимул минус стандартный стимул) в сериях 400-200 , 300-200 и 250-200 установлено, что амплитуда негативных различий в фронтальных, центральных и затылочных отведениях на участке 270-330 мс от начала стимула была статистически значимо больше в сериях 300-200 и 250-200 , чем в серии 400-200 , (p<0.05). Наиболее ранние (220 мс) значимые негативные различия возникали в серии 250-200 , затем - (270 мс) в серии 300-200 , и наиболее поздние - (650 мс) в серии 400-200 . В серии 400-200 в фронтальных и центральных отведениях с 200 мс от начала стимула даже наблюдалась статистически незначимая Различия ССПМ на стандартный и девиантный стимулы при отсутствии внимания к зрительным стимулам. Анализ ССПМ показал, что статистически значимый негативный сдвиг (1 - 4 мкВ) на девиантный стимул (РВ - разностная волна) с 200 до 700 мс от начала стимула наблюдался только в сериях, где длительность девиантного стимула меньше длительности стандартного ( 400-200 , 300- Активное внимание
Взаимодействие факторов внимание и длительность стимула F(3, 21 )=7.2Э7Э, р=.0015В 400 лс 300 лс 250 лс 150 лс 800 мс 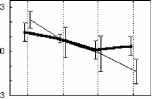 Отсутствие внимания 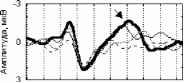 400 300 250 150 Длительность, мс отсутствие внимания - активное внимание 400 лс 300 лс 250 лс 150 лс 800 мс Рис. 3. Справа: ССПМ на стандартные стимулы разной длительности в условиях привлечения внимания и в его отсутствие в отведении СЗ. Стрелкой показан негативный компонент, зависящий от длительности стимула. Справа: Зависимость этого компонента от длительности стимула и внимания. установлено, что на пиковую амплитуду УП с высоким уровнем значимости влияет фактор длительности стимула (ANOVA Chi Sqr. (N = 10, df = 3) = 24.00000 p < .00002): амплитуда УП увеличивалась с ростом длительности стимула. Влияние внимания на амплитуду УП оказалось незначимым (F(1, 8)=1.3285, p=0.3). Влияние величины стимульного отклонения по длительности на характеристики ССПМ на девиантный стимул в отсутствие внимания. При сравнении ССПМ на девиантный стимул длительностью 200 мс в разных сериях при поточечном сравнении с помощью рангового дисперсионного анализа выявилось 3 промежутка, на которых обнаружено значимое влияние фактора величины стимульного отклонения правом фронтальном, правом центральном и теменных областях (рис 4). позитивная РВ на девиантный стимул (0.5-2 мкВ). Между сериями 100-200 и 150-200 различий по РВ на участке 150 - 400 мс не обнаружено. Значимые различия наблюдаются на участке 510-650 мс в фронтальных отведениях. Разностные ССПМ в серии 100-200 на этом участке были более позитивны, чем в серии 150-200 , что, вероятно, связано с появлением компонента Р3а, включающим механизм непроизвольного внимания [5]. Зависимость характеристик ССПМ от длительности стимула. При сравнении ССПМ на стандартные стимулы разной длительности установлено, что на участке от 150 до 500 мс от начала стимула как в условиях внимания к стимулам, так и в его отсутствие, наблюдается негативный сдвиг потенциала, максимально выраженный во фронтально-центральных областях и зависящий от длительности предъявляемого зрительного стимула (Рис.3). Вероятно, этот компонент представляет собой устойчивый потенциал (УП) [9]. С помощью дисперсионного анализа На участке 320-365 мс от начала стимула в значения амплитуд ССПМ были позитивными для серий 100-200 и 400-200 и негативными для остальных серий (p<0.05 - p<0.01). На участке 540600 мс эти различия сохранились, кроме серии 250-200 , для которой амплитуда стала равной нулю (p<0.05 - p<0.01). На участке 670-745 мс наиболее позитивными оказались ССПМ в серии 100-200 (p<0.05), что, вероятно, связано с включением механизма непроизвольного внимания. Влияние величины стимульного отклонения на характеристики ССПМ при активном внимании к длительности стимула (рис. 5). Известно, что компонент процессная 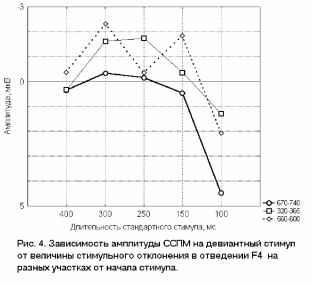 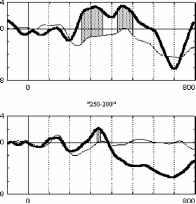 Разносшык СС1Ш (дет&ншый Е успоЕнях внимания - деЕнзншын Е успоЕнян игнорирования) 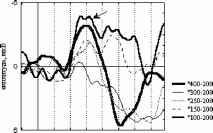 ЗаЖКНИОСГЬ ШППШУ7СЕ1 ПН VI Е4ЛЕНШЫ ЛНИ/ЛЬНОГО отклонения и обгаст отЕедення: FY.12, 84)=2.08В, р=0.02607  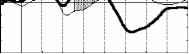 внимание - отсутствие внимания  100 50 -50 -О- С -100 -* р А- 0 Рис.5. Зависимость амплитуды ПН от величины стимульного отклонения. Слева: Среднегрупповые ССПМ на целевой девиантный стимул 200 мс в условиях внимания и на тот же стимул в условиях отсутствия внимания в отведении F4 при длительности нецелевого стандартного стимула 400, 250 и 100 мс. Справа вверху: Разностные ССПМ на целевой девиантный стимул в условиях внимания минус ССПМ на тот же стимул в условиях игнорирования при разных временных различиях между целевым девиантный и нецелевым стандартным стимулами в отведении F4. Стрелкой показана ПН. Справа внизу: Зависимость амплитуды ПН от величины стимульного отклонения и области отведения. негативность (ПН) может быть получен вычитанием из ССПМ на целевой релевантный стимул ССПМ на такой же стимул в условиях игнорирования [5]. При этом различия на участке 100-200 мс могут быть отнесены за счет усиления компонента N1 в условиях внимания, различия на участке 200-400 мс - за счет ПН и различия на участке от 400 мс - за счет РЗб на релевантный стимул [5]. При сравнении ССПМ на девиантный стимул в условиях привлечения внимания к стимулам и при невнимании к стимулам различия, связанные с появлением негативного компонента в условиях внимания на участке 200-400 мс от начала стимула, наблюдались при всех значениях стимульного отклонения во фронтальных и центральных отведениях. Максимальной амплитуды этот компонент достигал на 300 мс от начала стимула во фронтальных отведениях. Вычитанием ССПМ на девиантный стимул в условиях игнорирования из ССПМ на тот же стимул в условиях внимания были полученные разностные кривые и на участке 200-400 мс выделен компонент ПН. С помощью дисперсионного анализа установлено значимое взаимодействие факторов величины стимульного отклонения и области отведения (F(12, 84)=2.089, p=0.02607), связанное с тем, что во фронтальных, центральных и теменных областях амплитуда ПН была больше при большем стимульном отклонении (серии 400-200 , 100-200 ). Влияние фактора межполушарной асимметрии на амплитуду этого компонента оказалось незначимым. Обсуждение результатов Согласно полученным нами результатам, режимы 100-200 и 400-200 уже являются пограничными, поскольку эти длительности распознают только 39 и 64% испытуемых и для них ЭУ отличалась от ЭУп только согласно Т-критерию. Оказалось, что пороги различия длительности стимулов зависят от знака стимульного отклонения , при этом уменьшение длительности стандартного зрительного стимула по отношению к длительности девиантного распознается лучше, чем превышение длительности того же стимула над длительностью девиантного (см. Табл.1., режимы 300-200 и 100-200 ). По-видимому, зрительный аналог НР в чистом виде мог наблюдаться на временном промежутке с 200 мс от начала стимула в условиях отсутствия внимания к стимулам и получаться вычитанием ССПМ на стандартный стимул из ССПМ на девиантный стимул. При внимании к длительности стимула к НР должен был добавиться компонент процессная негативность (ПН), связанный со сличением стимула с эталоном, и, таким образом, амплитуда различий между стандартным и девиантным стимулом должна была увеличиться. Анализ литературы показал, что существование зрительной негативности рассогласования до сих пор убедительно не доказано. Существует относительно небольшое число работ, в которых непосредственно анализируется процесс сравнения зрительной информации. Среди них есть работы, в которых этот компонент был обнаружен [6], но есть также работы, в которых его обнаружить не удалось [7]. Так, в работе В. Б. Нечаева и др. (2000) был выделен затылочно-височный разностный компонент с латентным периодом 180-210мс, появляющийся на редкое повторение или изменение вербальных стимулов (букв русского алфавита) [6]. Этот компонент имел негативный максимум в затылочно-височных областях и позитивный - во фронтальных. В работе Woods D.L. и др. (1992) в ССПМ на девиантный зрительный стимул, появляющийся в правом или левом поле зрения, одно из которых было целевым, наблюдались бифазные изменения: положительная разностная волна с ЛП 100-130 мс в затылочных областях и негативная с ЛП 120-320 мс в височных областях, контралатеральных полю, в котором появлялся стимул. Подобно слуховой НР, зрительная НР увеличивалась при внимании [11]. С другой стороны, в некоторых работах (Nevill. H.J., Lawson D., 1987) не удалось обнаружить для зрительной модальности компонент, подобный слуховой НР [7]. В исследованиях Czigler I., Csibra G. (1990) негативность на девиантные стимулы, сходная со слуховой НР, появлялась только в ситуации внимания другим признакам стимула, являющимся целевыми [9]. В случае отсутствия внимания зрительная НР не наблюдалась. Как нами было установлено, значимая НР в отсутствии внимания наблюдалась только в сериях, где длительность девиантного стимула была меньше длительности стандартного (серии 400-200 , 300-200 , 250-200 ). В остальных сериях, даже при различиях, включающих непроизвольное внимание к девиантному стимулу (серия 100-200 ), значимой НР не обнаружено. В условиях внимания к стимулам негативная РВ на девиантный стимул увеличивается и становится значимой для всех серий, однако, в этом случае нельзя с уверенностью судить о наличии зрительной НР, поскольку РВ в задачах на селективное внимание является суммой НР на девиантный и ПН на целевой стимул [5]. Неожиданной оказалась зависимость НР от величины стимульного отклонения: при большем стимульном отклонении наблюдалась НР меньшей амплитуды. Согласно данным литературы, полученным для слуховых стимулов, амплитуда НР в большем числе работ увеличивается с ростом стимульного отклонения. Однако, Naatanen R, Gaillard A.W.K. (1983) указывают на то, амплитуда слуховой НР не может служить надежным коррелятом величины стимульного отклонения. Было установлено, что даже недетектируемый стимул может вызывать НР примерно той же самой величины, что и детектируемый [12]. Причина полученной нами обратной зависимости НР от величины стимульного отклонения легко объясняется, если проанализировать зависимость ССПМ на стандартный стимул от длительности стимула. Так, при увеличении длительности стимула на участке от 200 до 500 мс появляется негативный компонент, амплитуда которого увеличивается пропорционально длительности стимула. По-видимому, этот компонент представляет собой устойчивый потенциал (УП) [10]. Появление УП обычно связывают со стимулами длительностью более 600 мс [19], однако, УП зарегистрирован и более короткие стимулы [20]. Амплитуда УП увеличивается с ростом длительности стимула [19], а его ЛП составляет 150 мс [21]. В нашем случае амплитуда УП также растет с увеличением длительности стимула. Вероятно, УП на стандартные стимулы большей длительности и НР на девиантный стимул меньшей длительности характеризуются одинаковой латентностью (серии 300-200 , 250-200 ). Этот факт, объясняет полученную нами обратной зависимости амплитуды РВ от величины стимульного отклонения. Однако, сравнение ССПМ на девиантный стимул в отсутствие внимания в разных сериях показало, что зависимость амплитуды НР от величины стимульного отклонения все же не является прямо пропорциональной: при большем стимульном отклонении (серии 400-200 и 100-200 ) амплитуда ССПМ была менее негативна, чем в сериях с отклонением, пограничным для распознавания ( 300-200 и 150-200 ). Анализ изменений ССПМ, связанных с вниманием к длительности стимула (компонент процессная негативность , ПН) показал, что увеличение различий между нецелевым стандартным и целевым девиантным стимулами приводит к увеличению амплитуды этого компонента, что согласуется с литературными данными, полученными в условиях внимания к цвету, ориентации, формы зрительных стимулов и других атрибутов стимула, не связанных с пространственным положением [21, 5]. Выводы. 1. Порог различения длительности зрительных сигналов составляет около 100 мс и зависит от величины стимульного отклонения : уменьшение длительности зрительного стимула по сравнению с эталоном опознается лучше, чем его удлинение. 2. В отсутствии внимания к зрительным стимулам на участке от 200 до 500 мс после начала стимула обнаружен зрительный аналог НР. Зависимость зрительной НР от величины стимульного отклонения не является прямо пропорциональной, наибольшая НР возникает при низких значениях этого отклонения (50 мс), что указывает на возможность более тонкого различения длительности зрительных стимулов при автоматическом характере обработки информации. 3. Внимание к стимулам вызывает компонент ПН, связанный со сличением стимула с эталоном. Амплитуда этого компонента растет с увеличением временных различий между целевым и нецелевым стимулами. 4. Стимулы длительностью 150-400 мс вызывают УП на участке от 200 до 500 мс от начала стимула. Амплитуда этого компонента растет при увеличении длительности стимула. Настоящее исследование выполнено при финансовой поддержке гранта Конкурсного Центра Фундаментальных Исследований PD 02-1.4-433. Литература 1. Фонсова Н.А., Шестова И.А. Восприятие околосекундных интервалов времени Биологические науки. - 1988.- №3.- С.59-72. 1 2 |
|
| |||