
|
| |||
|
Разделы
Главная
Сапромат
Моделирование
Взаимодействие
Методы
Инновации
Индукция
Исследования
Факторизация
Частоты
Популярное
Как составляется проект слаботочных сетей?
Как защитить объект?
Слаботочные системы в проекте «Умный дом»
Какой дом надежнее: каркасный или брусовой?
Как правильно создавать слаботочные системы?
Что такое энергоэффективные дома?
|
Главная » Изменение Изменение с высотой над уровнем моря фауны дневных чешуекрылых (lepidoptera, diurna) на массиве горы Облачная (Южное Приморье) Мартыненко А.Б. (andrewmartynenko@mail.ru) Дальневосточный государственный университет, г. Владивосток Сихотэ-Алинь, средняя высота которого всего около 500-700 метров над уровнем моря, состоит из целой системы горных цепей, вытянутых в направлении с северо-востока на юго-запад и разделенных между собой речными долинами. Отдельные вершины, главным образом на восточных водораздельных хребтах, в пределах Приморского края достигают высоты 1200-1500 м. Наивысшая из них - гора Облачная, лежащая в верховьях реки Уссури. Она поднимается над уровнем моря на 1854 м. Отличительной особенностью массива горы Облачная является выраженность вдоль гипсометрического профиля полного спектра высотных поясов и подпоясов, свойственных обозначенной горной стране от смешанных кедрово-широколиственных лесов у подножья до горной тундры на вершине [3, 4, 24]. Дополнительную практическую значимость инвентаризационным исследованиям в указанном районе придает проектирование здесь организации одного из первых на Дальнем Востоке национального парка Зов тигра [26]. Облачная с давних пор приковывала внимание энтомологов, изучавших на ее примере хорологию различных таксономических групп. Вопросы высотной поясности фауны чешуекрылых данной территории наиболее полно были рассмотрены в публикациях А.И. Куренцова [8-12], определенное внимание было уделено этому и в трудах недавних русско-японских экспедиций [27, 28]. А.И. Куренцов в своих многочисленных публикациях утверждает, что на обследованных им горных поднятиях Южного Сихотэ-Алиня, как и в его средней части, с увеличением высоты над уровнем моря фауна насекомых, в том числе и дневных чешуекрылых, меняется не только количественно (беднеет), но и качественно. Это выражается в смене с подъемом в горы нескольких типов фауны: приамурской, охотско-камчатской и высокогорной. Ранее мы уже говорили о том, что анализ видовых списков хр. Ливадийский, входящего в состав гор Пржевальского [17], не подтверждает сформулированный Алексеем Ивановичем тезис. Тем не менее, до последнего времени оставались вопросы относительно наиболее высокой, центральной части Южного Сихотэ-Алиня, куда входит и массив горы Облачная. Решению обозначенной задачи и посвящено настоящее сообщение. МАТЕРИАЛ И МЕТОДЫ В основу работы положены материалы и наблюдения, сделанные нами на юге Чучуевского района Приморского края на южном макросклоне массива горы Облачная (рис. 1), в июне-июле 1992, 1996 и 2004 годов. Дополнительно, для составления наиболее полной картины высотно-поясного распределения были привлечены и доступные литературные источники [8, 11, 27, 28]. Выделение высотно-поясных территориальных образований на горе Облачная делалось нами согласно системе принципов, разработанной для гор Сибири и бассейна Амура В.Б. Сочавой [25], основываясь на структурных критериях. Для обозначения отдельного типа высотного ландшафта, то есть горизонтальной части соответствующего высотного пояса в границах геобиома (или его части) мы, вслед за Э.Г. Матисом [22], используем термин альтибиом . Применение этого термина в данном случае оправдано, поскольку его этимология ясно указывает, что он обозначает высотную модификацию зонального биома (геобиома). Количество альтибиомов (то есть основных высотно-поясных выделов) определялось набором зональных типов растительности (ландшафтов), сменяющих друг друга вдоль гипсометрического профиля. Так, в Приморском крае представлены следующие альтибиомы: альпийский, субальпийский, горной тайги, горной подтайги и температных лесов. При необходимости непереходный альтибиом (в регионе это в полной мере относится к альтибиому горной тайги и частично к альтибиому температных лесов) может быть разделен на 3 субальтибиома (верхний с признаками вышележащего альтибиома, средний - типичный и нижний с признаками нижележащего альтибиома), а переходный (горной подтайги и субальпийский) - на два: верхний с преобладанием черт вышележащего альтибиома и нижний с преобладанием черт нижележащего альтибиома. 135° 140° 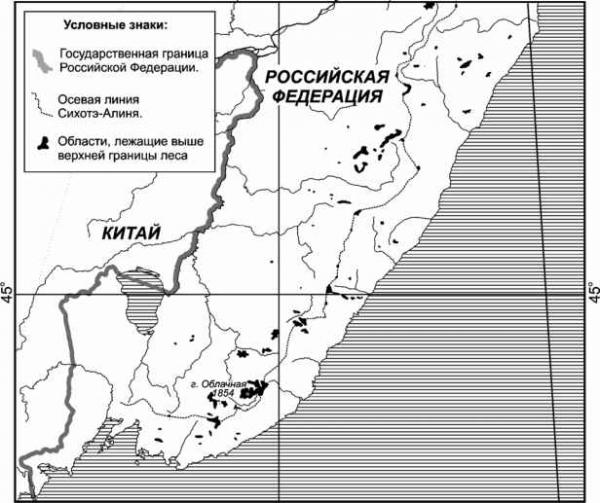 135° 140° Рис. 1. Местоположение массива горы Облачная (1854 м над ур. м) в Южном Сихотэ-Алине. Нумерация высотно-поясных выделов сверху вниз, а не снижу вверх, как это обычно делается, обусловлена соблюдением соответствия их номенклатуре широтно-поясных выделов, нумеруемых в Северном полушарии с севера на юг, то есть вдоль вектора увеличения теплообеспеченности [2]. При таком подходе в пределах гипсометрического профиля горы Облачная выделяется четыре альтибиома: у- гольцовый, 8 - подгольцовый, s - горнотаежный (субальтибиомы: sj - горной тайги редуцированного развития, S2 - горной тайги ограниченного развития и sj - горной тайги оптимального развития), Z -подтаежный, ) - температный (: nj - субальтибиом субнеморальных лесов). Для горы Облачная рассчитывалось изменение уровня видового богатства с высотой (А5100), показывающее на сколько видов в среднем изменяется (как правило убывает) видовое богатство вдоль гипсометрического профиля при подъеме на 100 м над ур. м. Также рассчитывался гипсометрический индекс (GAS ), представляющий собой отношение изменения уровня видового богатства при подъеме на 100 м к общему уровню видового богатства вдоль всего гипсометрического профиля [15, 16]. Типология ареалов строилась по эколого-ландшафтному критерию на основе локализации оптимума распространения относительно системы широтных и высотных поясов, долготных секторов [13, 14, 16]. При этом были учтены предложения по классификации и номенклатуре ареалов А.Ф. Емельянова [2]. Подразделение Палеарктики на подобласти принимается согласно районирования Л. А. Зенкевича (цит. по Негробову [23]), впоследствии уточненного А.Г. Вороновым с соавторами [1]. Номенклатура приводится в основном по справочнику Ю. П. Коршунова и П.Ю. Горбунова [7], впоследствии неоднократно дополнявшемуся первым соавтором [5, 6]. РЕЗУЛЬТАТЫ Всего в районе г. Облачная было отмечено 111 видов дневных бабочек, распределение которых по 4 альтибиомам (6 субальтибиомам) рассматривается ниже (табл. 1). Температный альтибиом (n) представлен только субнеморальным субальтибиомом. Субнеморальный субальтибиом (щ) представлен в рассматриваемом регионе в виде узкой полоски вдоль верховьев р. Уссури и на прилегающих сопках (500-650 м н.у.м.). В состав соответствующей ассамблеи Diurna входит 88 видов. В отличие от одноименной ассамблеи юго-западной части гор Пржевальского она беднее, что связано в первую очередь с отсутствием целого ряда температных видов (в частности Paramidea scolymus, Ninguta schrenckii, Ladoga moltrechti, Japonica saepestriata и Favonius taxila) и присутствием некоторых бореальных (в частности, Colias palaeno, Lopinga deidamia и Eumedonia eumedon). Альтибиом горных подтаежных лесов (Z) охватывает нижнюю часть склонов массива и низовья стекающих с него ключей (650-800 м н.у.м.). На субальтибиомы не подразделяется. Соответствующая альтибиому ассамблея Diurna, образованная 57 видами, принципиально мало отличается от аналогичной группировки юго-западной части гор Пржевальского. Различия между ними кроются в сравнительной бедности рассматриваемой ассамблеи и в ее обогащении бореальным элементом. Горно-таежный альтибиом (s) занимает наибольший высотный диапазон (800 -1500 м н.у.м.). В состав соответствующей ассамблеи Diurna входит 39 видов. Он подразделяется на 3 субальтибиома: горной тайги оптимального ()3), ограниченного ()2) и редуцированного (sj) развития. Субальтибиом горной тайги оптимального развития (s3) (800 - 1100 м н.у.м.). Растительность представлена преимущественно разнокустарниковыми елово-пихтовыми лесами с незначительным участием кедра корейского и ряда широколиственных пород. В этом наиболее богатом и типичном для южного Приморья таежном субальтибиоме было отмечено 33 вида Diurna. В этой ассамблее на фоне высокого удельного веса широко распространенных полизональных видов (Carterocephalus silvicola, Aporia crataegi, Lopinga achine, Nymphalis xanthomelas, Brenthis ino, Lycaenaphlaeas и др.) приблизительно в равной степени сочетаются представители таежной (Driopa eversmanni, Colias palaeno, Erebia ajanensis, Clossiana thore, Vacciniana optilete и др.) и неморальной фаун (Achillides maackii, Pieris dulcinea, Athymodes nycteis и др.). Субальтибиом горной тайги ограниченного развития (s2) (1000 - 1400 м н.у.м.) соответствует высокогорным ельникам с бедным флористическим составом. Участие пихты белокорой незначительно. Как и на юго-западе гор Пржевальского субальтибиом сильно затенен и крайне слабо населен дневными бабочками (отмечено только 20 видов). Соотношение фаунистических элементов то же, что и в предыдущей ассамблее. Большинство бабочек, как и в горной тайге оптимального развития, держатся вдоль тальвега ключей на освещенных участках (Carterocephalus palaemon, Pieris dulcinea, Erebia ligea и др.). Субальтибиом горной тайги редуцированного развития (si) (1400 - 1500 м н.у.м.) соответствует узкому поясу каменноберезовых кривоствольных лесов паркового характера с богатым травянистым ярусом, представленным преимущественно лесными видами. На отдельных участках в подлеске присутствует кедровый стланик. Ассамблея Diurna образована 26 видами, большинство из которых являются экологически пластичными и имеют широкий полизональный ареал. Самым предпочитаемым биотопом в этом субальтибиоме являются высокотравные луговины вблизи водосборных участков. Наиболее обычны в этих условиях Carterocephalus silvicola, Colias palaeno, Lopinga deidamia, Clossiana thore, Lycaena phlaeas и Eumedonia eumedon. Таблица 1 Видовое богатство и своеобразие высотно-поясных ассамблей Diurna на массиве горы Облачная
Сокращения. S - число видов; T - таксономическое своеобразие (по Смирнову); ареалогические группировки: AA - арктически-гипоарктические, гипоарктически-эвбореальные (аркто-альпийские) и альпийские, BM - эвбореальные, эвбореально-гемибореальные (бореомонтанные) и монтанные, HT - гемибореальные, гемибореально-температные и температные (CN - континентальные, PP -приокеанические, PS - полисекторные), TM - температные, температно-субтропические и тропико-субтропические (CN - континентальные, PP -приокеанические, PS - полисекторные), PB - внетропические полипоясные (CN -континентальные, PS - полисекторные). Субальпийский (или подгольцовый) альтибиом (8) (1500 - 1700 м н.у.м.) представлен, преимущественно, монодоминантными зарослями кедрового стланика, курамами, верещатнками и редкими разнотравными участками. На субальтибиомы не подразделяется. Ассамблея дневных чешуекрылых альтибиома небогата и как и предыдущая, в значительной мере складывается экологически пластичными видами (около 70% видового состава). В этом поясе в условиях каменистых обнажений с элементами разнотравья собирались Parnassius bremeri и Clossiana tritonia. Альпийский (или гольцовый) альтибиом (у) (более 1700 м н.у.м.) занимает самую верхнюю, водораздельную часть массива. На субальтибиомы не подразделяется. Преобладают тундровые ценозы, в понижениях на участках с хорошим почвенным увлажнением встречаются элементы альпийского разнотравья. В указанных условиях было отмечено всего 8 видов Diurna, что, к сожалению, не позволяет составить полное представление о его ассамблее. Выявляемости видов препятствуют не только частые пасмурные дни, но и постоянно дующие ветры. Как и в субальпийском альтибиоме обращает на себя внимание низкий удельный вес бореальных видов и полное отсутствие специфических альпийских на фоне преобладания экологически пластичных представителей. Относительно наиболее обычными бабочками были Pieris dulcinea, Lopinga deidamia, Aglais urticae и Nymphalis xanthomelas. В данных условиях изредка могут быть встречены самцы Driopa eversmanni, залетающие сюда с близлежащих ключей во время брачного лета. Постоянное обитание этого вида в условиях альпийского ландшафта южного Сихотэ-Алиня, вопреки предположению А.И. Куренцова [10-12], невозможно ввиду отсутствия кормового растения для гусениц (Corydalis gigantea). ОБСУЖДЕНИЕ Степень заселенности дневными бабочками различных альтибиомов (субальтибиомов) явно неоднородна. Как и следовало ожидать, наиболее богат всегда нижний субальтибиом, приуроченный к подножью гор и низкогорьям (табл. 1). Предсказуема также бедность чешуекрылыми средних и верхних горных поясов, получающих меньшее количество тепла и характеризующихся значительно меньшим набором биотопов (в частности постепенным выпадением долинных местоположений катены), бедной растительностью таежного, стланикового и горно-тундрового типов. Следует заметить, что на массиве горы Облачная таежные субальтибиомы населены дневными чешуекрылыми не богаче, чем подгольцовый и гольцовый, лежащие выше верхней границы леса. уровень сходства уровень включения: 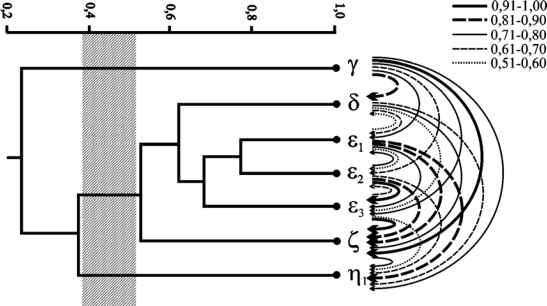 Рис. 2. Комбинированный граф сходства и включения субальтибиомных ассамблей Diurna на южном макросклоне горы Облачная. Номенклатуру высотно-поясных образований см. Материал и методика. Расчет изменения уровня видового богатства с высотой (ASioo) показал, что с подъемом в горы фауна заметно беднеет. При подъеме на 100 м по высоте видовое богатство снижается в среднем на 9,5 видов, что составляет 8,6% фауны. Еще сильнее беднеет фауна только на юге гор Пржевальского, где при подъеме на 100 м фауна теряет в среднем 14,1 видов, что составляет 6,9% от общего числа [17]. В средней и северной частях Сихотэ-Алиня за счет перераспределения видового богатства вдоль гипсометрического профиля темпы снижения с подъемом в горы ниже [15, 16, 18]. Еще меньше беднеет фауна Diurna с подъемом в горы западнее в субконтинентальных и континентальных районах Дальнего Востока и в Забайкалье [19-21]. Высокие темпы снижения видового богатства с увеличением гипсометрического уровня на горе Облачная обусловлены практически полным отсутствием характерной горной, бореомонтанной фауны Diurna, значительно богаче представленной в северной половине Дальнего Востока и в Забайкалье. Причем, наиболее резкое падение видового богатства происходит, как и в горах Пржевальского [17], на границе субнеморальных лесов с горными подтаежными лесами. Наиболее бедны темнохвойные леса, соответствующие горной тайге оптимального и ограниченного развития, где даже в горах Пржевальского было отмечено лишь 18 видов Diurna. Бедность горных елово-пихтовых лесов и ельников в данном случае, по-видимому, связана с тем, что большая часть этих лесов в силу сильной затененности и недостаточности трофического ресурса мало пригодна для заселения дневными бабочками. Выше в горах благодаря увеличению числа открытых биотопов наблюдается некоторое увеличение видового богатства. Сравнение альтибиомных ассамблей дневных чешуекрылых показало следующее (рис. 2). Все субальтибиомные ассамблеи группируются в три кластера: предгорный (ni), низкогорно-среднегорный (8 Sj - s3, ZZ и высокогорный (у). Предгорный кластер выделяется за счет высокого видового богатства и хорошей представленности температных приокеанических видов, низкогорно-среднегорный кластер характеризуется умеренным уровнем видового богатства и отсутствием многих южных видов, а высокогорный кластер выделяется за счет бедности видового состава, температные виды здесь не обнаружены. Граф включения, расположенный справа от дендрограммы, показывает, что высокогорные ассамблеи включаются в среднегорные, те - в низкогорные, а они, в свою очередь, в предгорные. Действительно, большинство горнотаежных видов, помимо соответствующего пояса, были нами отмечены в предгорье в условиях формации лиственничников. С другой стороны, часть температных приокеанических видов поднимаются в горы вплоть до верхней границы леса. Подтверждается это и расчетом общетаксономического своеобразия субальтибиомных ассамблей, с подъемом в горы оригинальность падает. Таким образом, данные А.И. Куренцова о наличии в Южном Сихотэ-Алине существенной качественной перестройки фауны [8-12] не подтверждается. На протяжении всего гипсометрического профиля относительно наиболее высоким уровнем видового богатства выделяются речные долины. Даже в горной темнохвойной тайге встречаются осветленные биотопы, наиболее благоприятные для заселения дневными бабочками. Сильнее всего с подъемом в горы беднеют биотопы горных водоразделов, последние уже в субнеморальных и подтаежных лесах населены отдельными видами Diurna. Вне зависимости от местоположения и высоты над уровнем моря наиболее заселены дневными чешуекрылыми частично осветленные древесно-кустарниковые биотопы, где присутствуют как лесные, так и луговые виды. ЗАКЛЮЧЕНИЕ Видовое богатство дневных чешуекрылых альтибиомов, в целом, сильно снижается с повышением гипсометрического уровня. Наиболее богаты субнеморальные леса, населенные 95 видами Diurna, а наиболее бедна горная тайга ограниченного развития, населенная только 18 видами. С подъемом в горы на 100 м по высоте видовое богатство снижается в среднем на 9,5 видов, что составляет 8,6% фауны. Падает и своеобразие фауны. Основной причиной этого является преобладание в фауне находящихся на северном пределе распространения восточноазиатских видов и практически полное отсутствие характерных таежных и гольцовых представителей. При этом существенной качественной перестройки фауны с подъемом в горы не происходит. ЛИТЕРАТУРА 1. Воронов А.Г. Дроздов Н.Н., Криволуцкий Д. А., Мяло Е.Г. Биогеография с основами экологии. М.: МГУ, 1999. 392 с. 2. Емельянов А.Ф. Предложения по классификации и номенклатуре ареалов Энтомол. обозр. 1974. Т.53. Вып.3. С.497-552. 3. Киселев А.Н., Кудрявцева Е.П. Высокогорная растительность Южного Приморья. М.: Наука, 1992. 117 с. 4. Колесников Б. П. Схема четвертичной истории высокогорной растительности Сихотэ-Алиня Проблемы ботаники. Вып. 12. Л.: Наука, 1974. С. 149-159. 5. Коршунов Ю. П. Дневные бабочки азиатской части России: Дополнения и исправления к книге. Новосибирск, 1996. 66 с. 6. Дневные бабочки азиатской части России: Новые описания и уточнения для книги. Новосибирск, 1998. 70 с. 7. Коршунов Ю.П, Горбунов П.Ю. Дневные бабочки азиатской части России: Справочник. Екатеринбург: Изд-во Урал. ун-та, 1995. 202 с. 8. Куренцов А.И. О вертикальном распределении чешуекрылых в Южном Сихотэ-Алине Зап. Влад. отд. геогр. об-ва. 1929. Т. 3. Вып. 20. С. 41-50. 9. Куренцов А. И. Дневные бабочки Приморского края. М: Главн. упр. по заповедникам, 1949. 119 с. 10. Куренцов А. И. Энтомофауна горных областей Дальнего Востока СССР (Эколого-географический очерк). М.: Наука, 1967. 96 с. 11. Куренцов А. И. Мои путешествия. Владивосток: Дальневост. кн. изд-во, 1973. 622 с. 12. Куренцов А.И. Зоогеография Дальнего Востока на примере распространения чешуекрылых - Rhopalocera. Л.: Наука, 1974. 160 с. 13. Мартыненко А. Б. Экология и распространение дневных чешуекрылых (Lepidoptera, Diurna) Приморского края. Автореферат диссертации на соискание ученой степени кандидата наук. Владивосток. 1998. 26 с. 14. Мартыненко А. Б. Вертикальное распределение булавоусых чешуекрылых (Lepidoptera, Diurna) на хребтах Воробей и Ливадийский Животный и растительный мир Дальнего Востока. Вып. 4. Уссурийск: УГПИ, 2000. С. 77-84. 15. Мартыненко А. Б. Альтибиомное и экотопическое распределение дневных бабочек (Lepidoptera, Diurna) в среднем Сихотэ-Алине Животный мир Дальнего Востока. Вып. 4. Благовещенск: БГПУ, 2002. С.157-166. 16. Мартыненко А.Б. Экология и география дневных чешуекрылых (Lepidoptera, Diurna) Приморского края. Владивосток: ДВГУ, 2004. 292 с. 17. Мартыненко А.Б. Влияние высотной поясности на структуру фауны дневных чешуекрылых (Lepidoptera, Diurna) в горах Пржевальского (Приморский край) Исследовано в России. 2005а. Т.8. С. 582-594. 18. Мартыненко А. Б. Высотная поясность фауны и биотопическое распределение дневных чешуекрылых в северном Сихотэ-Алине Бюлл. МОИП. Сер. биол. 2005б. (в печати). 19. Мартыненко А.Б. Высотно-поясное и биотопическое распределение дневных чешуекрылых (Lepidoptera, Diurna) на хр. Тункинские гольцы (Восточный Саян) Вестник БГУ, 2005в (в печати). 20. Мартыненко А. Б., Глущенко Ю. Н. Альтибиомная и биотопическая структура фауны дневных чешуекрылых (Lepidoptera, Diurna) в условиях гемибореальной лесостепи (на примере хр. Хамар-Дабан) Чтения памяти Алексея Ивановича Куренцова. Вып. 12. Владивосток: Дальнаука, 2002. С. 50-83. 21. Мартыненко А. Б. Чуркин С. В. Альтибиомно-биотопическое распределение дневных бабочек (Lepidoptera, Diurna) на северо-востоке Станового нагорья Животный мир Дальнего Востока. Вып. 4. Благовещенск: БГПУ, 2002. С. 167-192. 22. Матис Э. Г. Насекомые азиатской Берингии (принципы и опыт эколого-геосистемного изучения). М.: Наука, 1984. 312 с. 23. Негробов О.П. Краткий справочник по зоологической систематике. Воронеж: ВГУ, 1988. 112 с. 24. Пономаренко В. М. Верхняя граница леса на горе Облачной в Южном Сихотэ-Алине Сообщ. ДВФ АН СССР. 1960. Вып. 13. С. 73-77. 25. Сочава В.Б. Географические аспекты сибирской тайги. Новосибирск: Наука, 1980. 256 с. 26. Darman Yu., Karakin V., Martynenko A., Williams L. Conservation Action Plan for the Russian Far East Ecoregion complex. Part 1. Biodiversity and socio-economic assessment. Vladivostok-Khabarovsk-Blagoveshensk-Birobidzhan: WWF, 2003. 176 p. 27. Tashita M., Tshistjakov Y.A., Ono A. The diversity of butterfly communities in Southern Primorye Trans. lepid. Soc. Japan. 1997. Vol. 48. No. 3. P. 171-187. (in Japanese). 28. Takahashi M, Abe A., Abe R., Amano I., Yodoe K. A list of butterflies (Lepidoptera, Rho-palocera) collected in Southern Primorye in 1993 Far East. Entomol. 1996. No. 26. P.1- | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| |||